Napisane przez Zespół Naukowy Qubic
 Neuraxon Inteligencja Akademia — Tom 3
Neuraxon Inteligencja Akademia — Tom 3

1. Neuromodulacja w Mózgu: Podstawa Inteligencji Adaptacyjnej
Neuromodulacja odnosi się do zestawu mechanizmów, które regulują, jak system nerwowy funkcjonuje w danym momencie, nie zmieniając jego podstawowej architektury. Dzięki neuromodulacji mózg może uczyć się szybko lub wolno, stać się eksploracyjny lub konserwatywny i pozostawać otwarty na nowości lub skupiać się na tym, co już jest znane. Okablowanie się nie zmienia; to, co się zmienia, to sposób, w jaki to okablowanie jest używane. Ta koncepcja jest kluczowa dla zrozumienia inspirowanego mózgiem AI oraz architektury stojącej za Neuraxonem Qubic.
Receptory jonotropowe vs. metabotropowe: dwa czasowe skale sygnalizacji neuronowej
Aby właściwie zrozumieć neuromodulację, istotne jest rozróżnienie między dwoma formami działania chemicznego w mózgu. Z jednej strony są neuroprzekaźniki, które działają na receptory jonotropowe, takie jak glutaminian i GABA. Te receptory to kanały jonowe: gdy są aktywowane, powodują natychmiastowe zmiany elektryczne w neuronie, na poziomie milisekund. To odpowiada szybkiemu poziomowi obliczeń neuronowych: konkretne informacje są przesyłane, sygnały sensoryczne są integrowane, podejmowane są szybkie decyzje, a aktywność neuronalna, która podtrzymuje percepcję, ruch i myślenie w czasie rzeczywistym, jest generowana.
Z drugiej strony są neuroprzekaźniki, takie jak dopamina, noradrenalina, serotonina i acetylocholina, których główne działanie wywiera się przez receptory metabotropowe. Te receptory nie generują bezpośrednio sygnału elektrycznego. Zamiast tego aktywują wewnętrzne kaskady sygnalizacyjne, które modyfikują wewnętrzne właściwości neuronu przez dłuższe okresy czasu, sekundy, minuty lub dłużej. To reprezentuje powolny dynamiczny poziom przetwarzania neuronowego, który jest fundamentalny dla tego, jak mózg przystosowuje się i uczy.
Intuicyjny sposób myślenia o tej różnicy to metafora portu morskiego. Receptory jonotropowe są jak pływacy, surferzy lub małe łodzie, które szybko wchodzą i wychodzą. Receptory metabotropowe, w przeciwieństwie do tego, są jak duże statki towarowe. Aby mogły zacumować, potrzebne są pozwolenia, wymagana jest koordynacja, a logistyka portu musi być dostosowana. Te receptory metabotropowe zmieniają plastyczność synaptyczną i łatwość, z jaką neuron reaguje - ta wolna modulacja nie przesyła informacji, ale zamiast tego modyfikuje wewnętrzne zasady systemu.
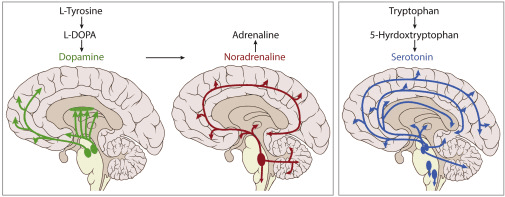
Cztery neuromodulatory: dopamina, noradrenalina, serotonina i acetylocholina
To tutaj wchodzą w grę główne systemy neuromodulacyjne. Każdy z tych czterech neuroprzekaźników odgrywa odrębną rolę w regulowaniu, jak mózg przetwarza informacje, uczy się i przystosowuje:
Dopamina, pochodząca głównie z obszaru brzusznego nakrywki i istoty czarnej, nie sygnalizuje przyjemności jako takiej, ale raczej, gdy coś jest istotne dla nauki. Dostosowuje wrażliwość systemu na błędy i nowość. Jak wykazał Schultz (2016) w swoim podstawowym badaniu nad kodowaniem błędów przewidywania nagrody dopaminy, dopamina sygnalizuje różnicę między oczekiwanymi a rzeczywistymi wynikami, mechanizm krytyczny dla uczenia się wzmocnionego zarówno w systemach biologicznych, jak i sztucznych.
Noradrenalina (norepinefryna), uwalniana głównie z miejsca sinawatego, reguluje pobudzenie i równowagę między eksploracją a eksploatacją. Gdy jej ton jest wysoki, mózg staje się bardziej wrażliwy na niespodziewane zmiany i mniej przywiązany do rutyn. To jest zgodne z integracyjną teorią zaproponowaną przez Aston-Jonesa i Cohena (2005), która łączy funkcję miejsca sinawatego-norepinefryny z kontrolą zysku adaptacyjnego i podejmowaniem decyzji w warunkach niepewności.
Serotonina, pochodząca z jąder szwu, moduluje nastrój, sen, hamowanie i stabilność behawioralną. Jak zbadano w pracy Dayana i Huysa (2009), serotonina nie zmusza systemu do szybkiego uczenia się, ale raczej do czekania, unikania impulsywnych reakcji i utrzymywania zachowania, gdy środowisko jest niepewne. Odtwarza krytyczną rolę w cierpliwości i długoterminowym planowaniu.
Acetylocholina, uwalniana z jąder podstawowych w mózgu, odgrywa centralną rolę w uwadze i uczeniu się zależnym od kontekstu. Ułatwia otwieranie sieci korowych na odpowiednie informacje sensoryczne i umożliwia plastyczność synaptyczną, gdy środowisko tego wymaga. Jest szczególnie ważna, gdy coś nowego musi być nauczone, co czyni ją niezbędną dla adaptacyjnych obliczeń neuronowych.
Dzięki temu połączonemu działaniu ten sam bodziec może powodować różne odpowiedzi w zależności od stanu neuromodulacyjnego. Obwód jest ten sam, ale sposób jego działania się zmienił. Dlatego mózg nie reaguje w ten sam sposób, gdy jest skupiony, jak gdy jest zmęczony, ani nie uczy się w ten sam sposób w rutynowych sytuacjach, jak w obliczu nowości lub zaskoczenia.
Poziom meta: okna plastyczności i adaptacyjne uczenie się
Istnieje również trzeci, głębszy poziom, który można zrozumieć jako poziom meta regulacji neuronowej. Ten poziom nie reguluje bezpośrednio aktywności neuronalnej ani jej prędkości, ale raczej warunki, w jakich system może zmienić się w sposób trwały. W mózgu zbieg aktywności między neuronami nie gwarantuje uczenia się. Aby połączenie mogło się wzmocnić lub osłabić, stan neuromodulacyjny musi na to pozwolić. To tak, jakby istniał cichy sygnał mówiący: „teraz tak” lub „teraz nie”.
Neuromodulacja działa zatem jako system, który otwiera lub zamyka okna plastyczności, decydując, kiedy błąd, doświadczenie lub zbieg okoliczności zasługuje na skonsolidowanie. Ta wieloskalowa architektura, szybka, wolna i meta, istnieje, ponieważ inteligentny system nie zawsze może stosować te same zasady. Jak wyjaśniła Marder (2012) w swojej przełomowej recenzji, neuromodulacja okręgów neuronowych to sposób, w jaki mózg osiąga elastyczność behawioralną bez odbudowywania swojej architektury.
Stan ciała, poziom energii, zmęczenie lub ból są częścią wewnętrznego środowiska. Nowość, zagrożenie, okazja, powtarzalność lub przewidywalność są częścią zewnętrznego środowiska. Systemy neuromodulacyjne tłumaczą te warunki na funkcjonalne stany. Poprzez dopaminę, noradrenalinę, serotoninę i acetylocholinę, mózg ocenia, czy sytuacja zasługuje na naukę, czy wymagana jest ostrożność, czy lepsza jest eksploracja, czy zachowanie, oraz czy błąd jest informacyjny, czy jedynie szumem. Środowisko nie dyktuje bezpośrednio odpowiedzi, ale moduluje zasady, według których mózg reaguje. Ta zasada leży u podstaw tego, co Friston (2010) opisał jako zasadę swobodnej energii, zjednoczoną ramę sugerującą, że mózg nieustannie minimalizuje zaskoczenie poprzez adaptacyjne wewnętrzne modele.
2. Dlaczego duże modele językowe i architektury transformatorowe nie mają neuromodulacji
Duże modele językowe (LLM) i architektury oparte na Transformerach nie posiadają neuromodulacji. Chociaż przetwarzają długie sekwencje i osiągnęły niezwykłe wyniki w przetwarzaniu języka naturalnego, brakuje im systemu, który dynamicznie reguluje reżim działania modelu podczas wnioskowania.
Statyczny charakter systemów AI opartych na Transformerach
Uczenie się w LLM zachodzi podczas faz treningowych, które są całkowicie oddzielne od użycia. Wagi są dostosowywane poprzez propagację błędu, a po zakończeniu treningu model wchodzi w stan ustalony. Podczas wnioskowania nie ma plastyczności i żadnej trwałej zmiany w funkcji kontekstu. System nie decyduje, kiedy jest odpowiedni moment na uczenie się, a kiedy powinien się ustabilizować, ponieważ nie uczy się, gdy działa. To jest fundamentalne ograniczenie, które potwierdziły niedawne badania, LLM nie mają prawdziwych wewnętrznych modeli świata i zdolności do adaptacji w czasie rzeczywistym.
Niektóre podejścia inspirowane neuromodulacją próbują przybliżyć pewne efekty poprzez dostosowanie parametrów, takich jak tempo uczenia podczas treningu, aktywowanie lub dezaktywowanie podsieci, lub modulowanie funkcji aktywacji. Jednak są to jedynie zewnętrzne optymalizacje, a nie wewnętrzne systemy, które regulują aktywność i plastyczność w czasie rzeczywistym. Jak argumentowali Mei, Müller i Ramaswamy (2022) w Trends in Neurosciences, informowanie głębokich sieci neuronowych przez wieloskalowe zasady systemów neuromodulacyjnych pozostaje otwartym wyzwaniem, które obecne architektury LLM nie rozwiązały.
Chociaż neuromodulacja jest czasami wspominana w kontekście AI, LLM i Transformery pozostają częściowymi przybliżeniami, a nie systemami porównywalnymi z mózgiem. Różnica między statycznymi obliczeniami macierzowymi a dynamiczną, zależną od stanu regulacją występującą w biologicznych sieciach neuronowych jest dokładnie tym, co sprawia, że architektury AI inspirowane mózgiem, takie jak Neuraxon, są niezbędnym krokiem w kierunku adaptacyjnej inteligencji sztucznej.
3. Jak Neuraxon oblicza neuromodulację: architektura AI inspirowana mózgiem
W Neuraxon obliczenia są procesem, który rozwija się w czasie ciągłym. Kod wyraża system, który utrzymuje wewnętrzne stany, s(t), które ewoluują nawet w braku wyraźnych zewnętrznych bodźców. Stany te wpływają na przyszłe zachowanie, tworząc żywy system neuronowy, który jest zawsze aktywny, koncepcja ta jest szczegółowo badana w pracy badawczej Neuraxon.
Szybka, wolna i meta dynamika w obliczeniach neuronowych
Neuraxon wyraźnie uwzględnia szybkie, wolne i meta dynamiki, odzwierciedlając wieloskalową architekturę czasową występującą w biologicznym mózgu. Szybkie dynamiki rządzą natychmiastową propagacją aktywności, analogicznie do szybkiego sygnalizowania neuronalnego przez receptory jonotropowe. Wolne dynamiki wprowadzają akumulację, trwałość i stabilizację wzorców, pozwalając systemowi na zachowanie informacji poza chwilą, podobnie jak receptory metabotropowe modulują funkcję neuronową przez sekundy i minuty. Dynamika meta działa na zasady interakcji między tymi poprzednimi, modulując, kiedy system staje się bardziej wrażliwy na zmiany i kiedy ma tendencję do zachowania swojego stanu.
Neuromodulacja w Neuraxon nie jest wdrażana jako zewnętrzne dostosowanie parametrów. System nie decyduje wyraźnie, co ma się uczyć, ale raczej w jakich warunkach może się zmienić. To odzwierciedla sposób, w jaki biologiczne neuromodulatory, takie jak dopamina i serotonina, tworzą okna plastyczności, a nie bezpośrednio kodują informacje. Możesz na własne oczy zbadać te dynamiki za pomocą interaktywnej symulacji 3D Neuraxon na HuggingFace Spaces, gdzie możesz dostosować poziomy dopaminy, serotoniny, acetylocholiny i norepinefryny w czasie rzeczywistym i obserwować, jak wpływają one na zachowanie sieci.
Od zasad biologicznych do zdecentralizowanej AI
To podejście nie reprodukuje molekularnej ani anatomicznej złożoności mózgu, co obecnie jest niemożliwe do powtórzenia. Nie ma tysięcy receptorów ani prawdziwych biologicznych sieci. Jednak zachowuje i oblicza zasadniczą zasadę: inteligencja jest adaptacyjna, a zatem wymaga wewnętrznej dynamiki, stanu i modulacji.
Architektura neuromodulacji Neuraxon jest kluczową częścią szerszej wizji Qubic dotyczącej zdecentralizowanej AI. Integrując Neuraxon z ewolucyjnym frameworkiem Aigarth Intelligent Tissue, Qubic tworzy system, w którym miliony architektur opartych na Neuraxon mogą ewoluować, konkurować i poprawiać się poprzez rozproszone obliczenia, napędzane mechanizmem konsensusu Useful Proof of Work (UPoW) sieci Qubic.
4. Zbadaj neuromodulatory z interaktywnym demo Neuraxon
Chcesz doświadczyć, jak działa neuromodulacja w systemie AI inspirowanym mózgiem? Demo Neuraxon Mood Mixer pozwala dostosować poziomy dopaminy, serotoniny, acetylocholiny i norepinefryny w czasie rzeczywistym i obserwować, jak te neuromodulatory wpływają na zachowanie sieci neuronowej. To praktyczny sposób na zrozumienie zasad omówionych w tym artykule i zobaczenie różnicy między statycznym obliczeniem AI a dynamicznym, zależnym od stanu przetwarzaniem.
5. Matematyka stojąca za wieloskalową neuromodulacją Neuraxon
Dynamika czasowa w Neuraxon jest regulowana przez trzy równania różniczkowe, które uchwycają szybkie, wolne i meta czasowe skale obliczeń neuronowych:

Tutaj, τ_fast < τ_slow < τ_meta odzwierciedlają ich odrębne skale czasowe, przy czym τ_meta jest znacznie większe, aby uchwycić „ultraspowolny” charakter efektów metabotropowych. Ta ramka matematyczna bezpośrednio wdraża biologiczną zasadę, że neuromodulacja działa na znacznie wolniejszych skalach czasowych niż szybkie przesyłanie synaptyczne, jak opisano przez Northoffa i Huanga (2017) w ich pracy na temat tego, jak czasowe dynamiki mózgu mediują świadomość.
Bibliografia naukowa
Dayan, P., & Huys, Q. J. M. (2009). Serotonina, hamowanie i negatywny nastrój. PLoS Computational Biology.
Marder, E. (2012). Neuromodulacja okręgów neuronowych: powrót do przyszłości. Neuron.
Schultz, W. (2016). Kodowanie błędu przewidywania nagrody dopaminy. Dialogi w Neurologii Klinicznej.
Aston-Jones, G., & Cohen, J. D. (2005). Integracyjna teoria funkcji locus coeruleus-norepinefryny. Roczny przegląd neurologii.
Mei, L., Müller, E., & Ramaswamy, S. (2022). Informowanie głębokich sieci neuronowych przez wieloskalowe zasady systemów neuromodulacyjnych. Trends in Neurosciences.
Friston, K. (2010). Zasada swobodnej energii: zjednoczona teoria mózgu? Nature Reviews Neuroscience.
Northoff, G., & Huang, Z. (2017). Jak czas i przestrzeń mózgu mediują świadomość i jej zaburzenia? Świadomość i poznanie, 57, 1–10.